Die Herausforderung
von Hyojung Kang, Rachel Orlowsky und Gregory J. Gerling (Universität von Virginia)
Vorgestellt auf der Wintersimulationskonferenz 2017
Bei Säugetieren wird der Tastsinn durch in die Haut eingebettete sensorische Rezeptoren kodiert. Bei einer Klasse von Rezeptoren in der Maus folgt die Architektur der Merkelzellen, der nicht-myelinisierten Neuriten und der Heminoden bestimmten Erneuerungs- und Umbautrends über die Stadien des Haarzyklus im Alter von 4 bis 10 Wochen. Da es derzeit unmöglich ist, solche Trends über den Haarzyklus eines einzelnen Tieres zu beobachten, werden in dieser Arbeit diskrete Ereignissimulationen eingesetzt, um Strategien für die Dynamik der Merkelzellen und Heminoden zu identifizieren und zu bewerten. Die Ergebnisse stimmen gut mit den beobachteten Daten überein und zeigen, dass das Basismodell das dynamische Umbauverhalten zwischen den Stadien des Haarzyklus nachbildet - basierend auf bestimmten Hinzufügungs- und Entfernungsstrategien und geschätzten Wahrscheinlichkeiten, die an die Bestandteile von Merkelzellen, terminalen Zweigneuriten und Heminoden gebunden sind. Die Analyse zeigt außerdem, dass bestimmte Strategien einen größeren Einfluss haben als andere. Dieser Einsatz von Berechnungen ist ein neuartiger Ansatz zum Verständnis der neuronalen Entwicklung.
Einführung
Der Tastsinn ist der Schlüssel zu alltäglichen Verhaltensweisen wie Nahrungsaufnahme, soziale Bindungen und Vermeidung von körperlichen Schäden. Bei Säugetieren wird der Tastsinn durch sensorische Rezeptoren in der Haut kodiert (Kandel 2012). Zu den sensorischen Rezeptoren gehören Afferenzen für leichte Berührungen in der Haut sowie solche, die Informationen über Propriozeption, Chemorezeption und Schmerz signalisieren. Sowohl die sensorischen Rezeptoren als auch die Haut erneuern und modellieren sich ständig, um die Barriere im Normalzustand und nach Verletzungen aufrechtzuerhalten (Chung 2010; Marshall 2016; Müller-Röver 2001; Rajan 2003). In der behaarten Haut sind die Nervenendigungen der Merkelzellen in spezialisierten Epithelstrukturen, den sogenannten "Touch Domes", zusammengefasst (Plikus 2008). Mäuse haben Hunderte von Touch Domes in ihrer behaarten Haut, und auch der Mensch hat ähnliche, wenn auch subtil unterschiedliche Nervenendigungen. Allgemeiner ausgedrückt: Merkel-Zell-Rezeptoren sind sowohl in der behaarten als auch in der kahlen Haut von Säugetieren zu finden, wobei die lokalen Rezeptor- und Hautstrukturen in jedem Fall variieren. In rezeptiven Populationen helfen diese Afferenzen dabei, Informationen über die Ränder und die Krümmung von Reizen sowie andere Eigenschaften zu signalisieren (Johnson 2001).
Die Dynamik der Architektur des Merkelzell-Neuritenkomplexes ist gerade erst im Entstehen begriffen (siehe Abbildung 1 und Lesniak 2014). Es wurde beobachtet, dass die Verbindung einer Merkel-Zelle mit einem terminalen Neuriten oder die Entfernung von diesem sowie Neuriten von Heminodien Trends folgen, die für die Stadien des Haarzyklus bei der Maus spezifisch sind (Marshall 2016). Insbesondere werden bei Mäusen mit zunehmendem Alter mehrere synchronisierte Haarzyklen beobachtet, bei denen sich die Behaarung des Tieres am gesamten Körper wellenförmig verändert (Müller-Röver 2001). Der spontane Haarzyklus von Mäusen lässt sich in vier Phasen einteilen: Erstes Telogen: 4 Wochen, Anagen: 5-6 Wochen, Katagen: 6 Wochen, und zweites Telogen: 9-10 Wochen. Danach beginnt der Haarzyklus in eine Mosaikphase einzutreten, in der sich die Haare auf dem Körper des Tieres nicht wellenförmig verändern, sondern an scheinbar zufälligen Stellen auf der Hautoberfläche in unterschiedlicher Geschwindigkeit ausfallen und wieder nachwachsen. Kutane Neuronen und Merkel-Zellen können während der Regeneration der Haarfollikel an Plastizitätsmechanismen beteiligt sein; die Dynamik und die physiologischen Folgen der neuronalen Plastizität in den Berührungsrezeptoren sind jedoch nicht vollständig geklärt (Moll 1996; Nakafusa 2006).
Die Beobachtung des Umbaus von Haarbornen ist nur begrenzt möglich, da wir derzeit nicht in der Lage sind, spezifische Endorgane durch den Haarzyklus zu verfolgen. Ein Modellierungsansatz hingegen ermöglicht eine detaillierte Verfolgung der Endorgane und aller ihrer Komponenten durch alle Stadien des Haarzyklus. Darüber hinaus gibt es trotz des Einsatzes von Modellen der diskreten Ereignissimulation (DES) in der biologischen Forschung immer noch sehr wenige veröffentlichte Beispiele für umfassend validierte Modelle. Daher hat unsere Gruppe vor kurzem ein Computermodell entwickelt, um eine erste Reihe von Regeln für den Umbau von Sprossen zu testen (Marshall 2016). Bei diesem Top-Down-Ansatz wurden Populationsstatistiken aus beobachteten Daten verwendet, um das Rechenmodell zu erstellen. Unter Verwendung der Populationsstatistiken als Referenzpunkte wurden iterativ Endorgan-Bestandteile erstellt und gelöscht, um die beobachteten Daten widerzuspiegeln. Jeder der Übergänge zwischen den Stadien des Haarzyklus wurde als separate Simulation modelliert. Innerhalb jeder Simulation gab es eine Reihe von Iterationen, bei denen Merkel-Zellen und/oder Heminodien nach nur vier probabilistischen Regeln hinzugefügt oder entfernt werden konnten, die auf morphometrischen Daten basierten. Bei diesem Versuch wurden vier Strategien für die Dynamik von Merkelzellen und Heminoden aus Beobachtungsdaten ermittelt und bewertet.
Die Studie (Marshall 2016) trug zwar dazu bei, die Regeln zu erklären, die die Umbauprozesse zwischen Merkelzellen und Neuriten (oder Arbor) steuern, aber diese Berechnungsmodelle haben mehrere Einschränkungen. Zum Beispiel waren die einzelnen Endorganbestandteile von Merkelzellen, terminalen Neuriten und Heminodien zwischen den Phasen des Haarzyklus nicht nachvollziehbar. Die Beschränkung auf einen Top-Down-Ansatz behinderte zudem die Entwicklung von Strategien für die Umgestaltung der Adern, die sich auf einzelne Komponenten des Endorgans und nicht auf das Endorgan selbst beziehen. Außerdem war es aufgrund der Modellstruktur nicht einfach, Was-wäre-wenn-Szenarien zu testen und die optimalen Parameter für die maßgeblichen Regeln zu bestimmen. Schließlich gibt es in diesem Modell keine einfachen Optimierungs- oder Experimentierfunktionen, was das Testen neuer Parameter, die vorher nicht definiert wurden, einschränkt. Daher haben wir versucht, diese Einschränkungen durch den Einsatz eines DES-Ansatzes zu überwinden. Ziel dieser Studie ist es, zu zeigen, wie ein DES-Modellierungsansatz dazu beitragen kann, die zugrundeliegenden Mechanismen der Veränderungen in den Arboren im Verlauf des Haarzyklus zu verstehen. Im Gegensatz zu den früheren Bemühungen um eine computergestützte Modellierung verfolgt der hier vorgestellte Ansatz einen Bottom-up-Ansatz mit kodierten Wahrscheinlichkeitsregeln für jeden spezifischen Bestandteil, wenn das übergeordnete Endorgan den Haarzyklus durchläuft. Die Strategien sind eine Kombination aus bedingten und probabilistischen Regeln. Im Gegensatz zum Computermodell wurde jede Merkelzelle, jeder terminale Neuritenast und jede Heminode, die zu einem bestimmten Endorgan gehören, einzeln untersucht, und die Regeln für ihre Hinzufügung und Löschung spiegeln die Hypothesen über die biologischen Eigenschaften wider. Dieser Bottom-up-Ansatz, bei dem Endorgane, Heminoden und terminale Neuritenverzweigungen als unterschiedliche Entitäten modelliert werden, ermöglichte es uns, die regelnden Strategien zu verfeinern und optimale Parameter zu schätzen, die bei den Umbauprozessen eine Rolle spielen. Die Merkel-Zellen werden als ein Attribut der terminalen Äste modelliert, das die Richtlinien und Parameter beeinflusst.
Die Lösung
Methode
Ziel dieser Arbeit ist es, mit Hilfe von DES Prinzipien zu identifizieren, die festlegen, wie sich die Äste im Verlauf des Haarzyklus verändern, und Strategien für die Dynamik der Merkel-Zellen und der Heminoden zu bewerten. Es werden verschiedene Kombinationen von Geschwindigkeitsentwicklungen und Inkorporationsregeln simuliert, um Vorhersagen von Endorganarmen zum Vergleich mit spontanen Haarzyklusbeobachtungen zu erhalten.
Abstraktion der Biologie zur Beschreibung der modellierten Inputs, Outputs und Übergänge
Wir betrachten den Merkelzell-Neuritenkomplex in der behaarten Haut der Maus. Obwohl die Architektur des Merkelzell-Neuritenkomplexes in Bezug auf seine Bestandteile noch komplizierter sein kann, ist die in Abbildung 1 dargestellte Abstraktionsebene einigermaßen typisch und wird zur Beschreibung der Ein- und Ausgänge des hier zu erstellenden Modells verwendet.

Auf der Grundlage früherer Modellierungsbemühungen (Marshall 2016) wurde ein vorläufiger Satz von Regeln entwickelt, die in der vorliegenden Arbeit modifiziert wurden, um von einer Bottom-up-Perspektive auszugehen (siehe Tabelle 1). Im Allgemeinen überprüfen wir zuerst den Zustand eines Heminodus, indem wir die zugehörigen terminalen Äste und Merkelzellen überprüfen.
Diskrete Ereignissimulation
Modellbeschreibung
Es wurde ein DES-Modell erstellt, um die während der Übergänge im Haarzyklus auftretende Dynamik zu reproduzieren und die Schlüsselparameter zu bestimmen, die das dynamische Verhalten beeinflussen. Das Modell bestand aus drei Entitätstypen, von denen jeder Endorgane, Heminodien und Endverzweigungen repräsentiert. Merkelzellen wurden als binäres Attribut von Endzweig-Entitäten modelliert: Jeder Endzweig hat entweder eine Merkelzelle (1) oder keine Merkelzelle (0). Die Struktur der Äste wurde als eine Hierarchie aus zwei Chargenschichten modelliert. Jede Endorgan-Entität besteht aus einer Reihe von Heminode-Entitäten, und jede Heminode-Entität besteht aus einer Reihe von Endzweig-Entitäten. Während jedes Endorgan einen kompletten Haarzyklus durchläuft, ändert sich seine Zusammensetzung kontinuierlich durch Batching- und Unbatching-Prozesse mit unterschiedlichen Regeln. Das Modell wurde mit dem Simio Simulation and Scheduling Software Package erstellt.
Parameterabschätzung und Validierung
Zur Entwicklung des Basismodells wurden Beobachtungsdaten aus der vorangegangenen Studie verwendet (Marshall 2016). Zu den wichtigsten Parametern in First Telogen gehörten die Anzahl der Heminoden pro Endorgan, die Anzahl der terminalen Verzweigungen pro Endorgan und die Anzahl der Merkelzellen pro Endorgan, die Anzahl der terminalen Verzweigungen pro Heminode und die Anzahl der Merkelzellen pro Heminode. Diese Parameter wurden anhand empirischer Verteilungen geschätzt. Die Dauer jedes Prozesses betrug 4, 2 bzw. 4 Wochen zwischen den Stadien der Haarzyklen.
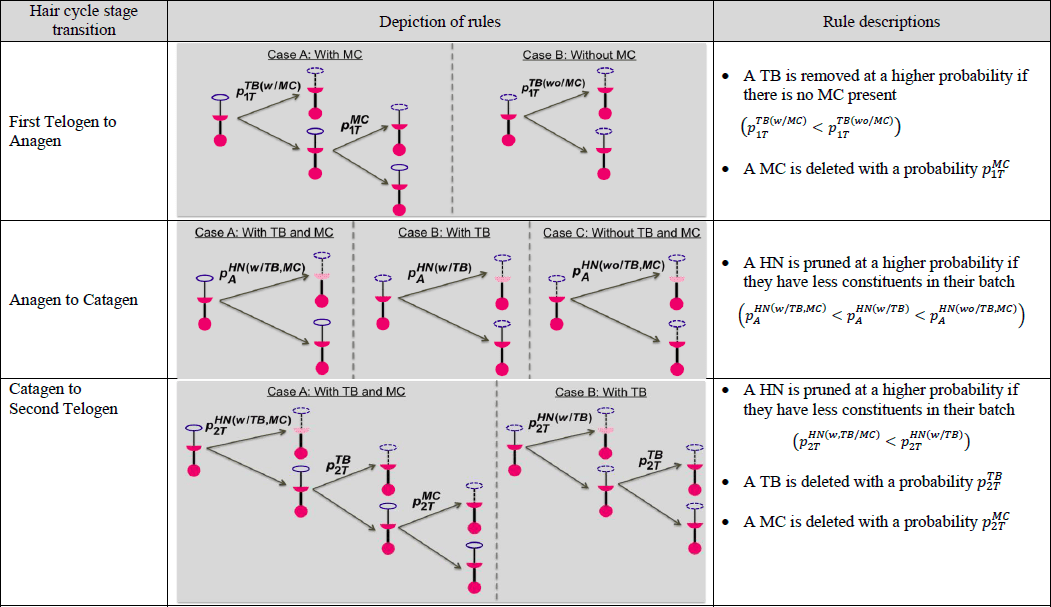
Tabelle 1. Grafische und textbasierte Darstellung der Regeln, die mit jedem der drei Übergänge des Haarzyklus verbunden sind, wobei MC = Merkel-Zelle und TB = Terminaler Zweig und![]() die Wahrscheinlichkeit darstellt, dass eine Komponente eines Endorgans x im Stadiums entfernt/beschnittenwird. xumfasst MC, TB und HN. Der Begriffs umfasst 1T (erstes Telogen), A (Anagen), C (Katagen) und 2T (zweites Telogen).
die Wahrscheinlichkeit darstellt, dass eine Komponente eines Endorgans x im Stadiums entfernt/beschnittenwird. xumfasst MC, TB und HN. Der Begriffs umfasst 1T (erstes Telogen), A (Anagen), C (Katagen) und 2T (zweites Telogen).
Eines der Ziele dieser Studie war die Schätzung der Wahrscheinlichkeiten für die Hinzufügung und Löschung jeder Merkel-Zelle, jedes terminalen Astes und jeder Heminode in Bezug auf ein Endorgan zwischen den Stadien. Zur Kalibrierung dieser unbekannten Modellparameter wurden drei Hauptergebnisse berücksichtigt: die durchschnittliche Anzahl von Heminoden pro Endorgan, die durchschnittliche Anzahl von Merkelzellen pro Endorgan und die durchschnittliche Anzahl von Merkelzellen pro Heminode. Die Kalibrierung des Modells wurde in jedem Stadium chronologisch durchgeführt, und zwar eines nach dem anderen, da die Wahrscheinlichkeiten in den verschiedenen Stadien unabhängig voneinander sind, die Populationsstatistiken der einzelnen Haarzyklusstadien jedoch vom vorhergehenden Stadium abhängig sind. Das kalibrierte Basismodell wurde mit 25 Wiederholungen durchgeführt und durch den Vergleich seiner Ergebnisse mit den drei Hauptergebnissen der Beobachtungsdaten validiert. Es wurde eine Sensitivitätsanalyse durchgeführt, um die Auswirkungen von Änderungen der Deletions-/Zusatzwahrscheinlichkeiten auf die Hauptergebnisse zwischen Katagen und zweitem Telogen zu verstehen. Wir wählten die beiden Stadien, weil während des letzten Übergangs des Haarzyklus mehr Umbauaktivitäten unter Beteiligung von Heminodien, terminalen Verzweigungen und Merkelzellen stattfinden als bei anderen Übergängen.
Die Auswirkungen auf das Geschäft
Ergebnisse
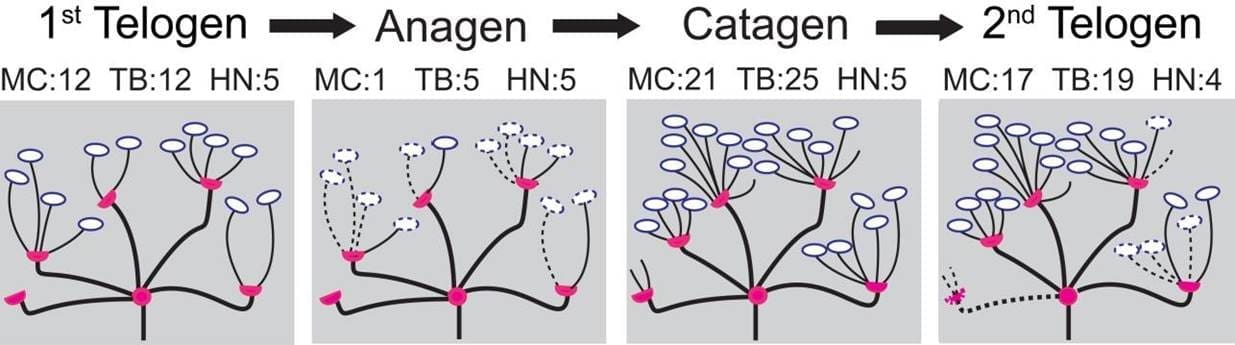
Eine Beispielsimulation eines Endorgans durch die vier Stadien des Haarzyklus ist in Abbildung 2 dargestellt. Auf der Grundlage der in der vorangegangenen Studie (Marshall 2016) entwickelten konzeptionellen Deletions-/Zusatzstrategien zwischen den Haarzyklen haben wir Bottom-up-Wahrscheinlichkeiten generiert, wie in Tabelle 2 (nächste Seite) dargestellt. Beim Übergang vom ersten Telogen zum Anagen beträgt die Wahrscheinlichkeit, dass ein terminaler Zweig entfernt wird, 0,9, wenn er keine Merkel-Zelle enthält, und 0,5, wenn eine Merkel-Zelle vorhanden ist. Während des Übergangs wird eine Merkel-Zelle mit einer Wahrscheinlichkeit von 0,9 entfernt. Beim Übergang von Anagen zu Katagen ist es wahrscheinlicher, dass Heminoden beschnitten werden, wenn ihre Bestandteile leer sind. So wird ein Heminode mit einer Wahrscheinlichkeit von 0,2 beschnitten, wenn er keine endständigen Äste, sondern Merkelzellen enthält, mit einer Wahrscheinlichkeit von 0,1, wenn er endständige Äste, aber keine Merkelzellen enthält, und mit einer Wahrscheinlichkeit von 0,05, wenn er sowohl endständige Äste als auch Merkelzellen enthält. Beim Übergang vom Katagen zum zweiten Telogen folgen die Heminknoten einer ähnlichen proportionalen Löschungspolitik. Während des Übergangs verlieren die Heminknoten mit einer Wahrscheinlichkeit von 0,8 mindestens einen terminalen Zweig. Die Anzahl der zu löschenden Endverzweigungen liegt zwischen 1 und 3 mit der gleichen Wahrscheinlichkeit. Eine Merkel-Zelle wird mit einer Wahrscheinlichkeit von 0,9 gelöscht.
Das Basissimulationsmodell wurde anhand der von Marshall et al. (2016) gesammelten Beobachtungsdaten validiert. Tabelle 3 vergleicht die aus 20 Beobachtungsdaten abgeleitete mittlere Anzahl von Merkelzellen und Heminodien in jedem Stadium mit den aus dem DES-Modell mit 20 Replikationen geschätzten Werten. Die Modellergebnisse waren den tatsächlichen Daten sehr ähnlich, was darauf hindeutet, dass das Basissimulationsmodell eine angemessene Darstellung des Arbor-Systems ist (Banks 2000).
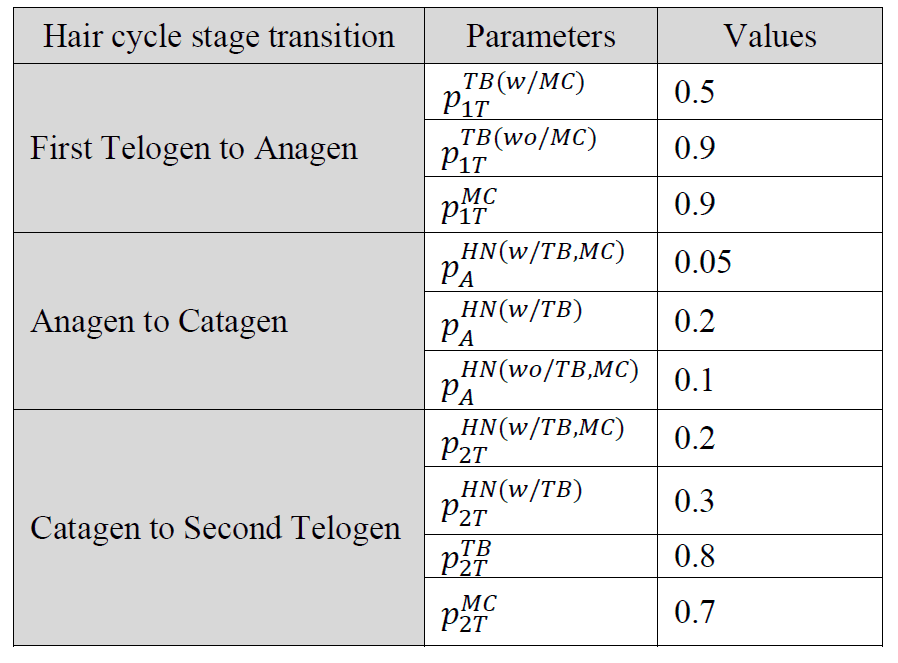
Tabelle 2. Parameterwerte, die im Simulationsmodell bei jedem der drei Übergänge des Haarzyklus verwendet werden, wobei MC = Merkel-Zelle und TB = terminaler Zweig und![]() die Wahrscheinlichkeit darstellt, dass eine Komponente eines Endorgans x in der Phases entfernt/beschnittenwird xumfasst MC, TB und HN. Der Begriffs umfasst 1T (erstes Telogen), A (Anagen), C (Katagen) und 2T (zweites Telogen).
die Wahrscheinlichkeit darstellt, dass eine Komponente eines Endorgans x in der Phases entfernt/beschnittenwird xumfasst MC, TB und HN. Der Begriffs umfasst 1T (erstes Telogen), A (Anagen), C (Katagen) und 2T (zweites Telogen).
|
Beobachtungsdaten (Mittelwert) |
Modellergebnisse (Mittelwert [95 % CI]) |
|||
|
Merkel-Zellen |
Heminodien |
Merkel-Zellen |
Heminodien |
|
| Erstes Telogen |
13 |
5 |
13 [12.4, 17.3] |
5 [5.3, 5.5] |
| Anagen |
0 |
5 |
0 [0, 0.7] |
5 [5.2, 5.5] |
| Katagen |
25 |
4 |
24 [24.1, 24.8] |
4 [3.9, 4.2] |
| Zweites Telogen |
15 |
3 |
14 [13.8, 14.2] |
3 [3.2, 3.9] |
Abbildung 3 vergleicht die Ergebnisse der beobachteten und simulierten Daten. Die Modelldynamik, die sich ausschließlich auf Hinweise im Zusammenhang mit dem Haarzyklus stützt, erzeugt Äste, die mit den experimentellen Beobachtungen gut übereinstimmen. Die Verteilung der Heminodien, terminalen Äste und Merkel-Zellen pro Endorgan bei jedem Haarzyklus und die Muster der Veränderungen waren zwischen den Beobachtungsdaten und den Simulationsergebnissen ähnlich. Das Simulationsmodell ermöglichte es uns auch, jedes Endorgan über den Verlauf des Haarzyklus zu verfolgen. So stellen beispielsweise die blauen und orangefarbenen Punkte in den Feldern D, E und F in Abbildung 3 jeweils zwei verschiedene Endorgane dar. Das dynamische Umbauverhalten zwischen den experimentellen Beobachtungen und den Modellergebnissen wurde jedoch nicht auf der Ebene der Endorgane verglichen, da die Beobachtungsdaten nicht nachvollziehbar waren.
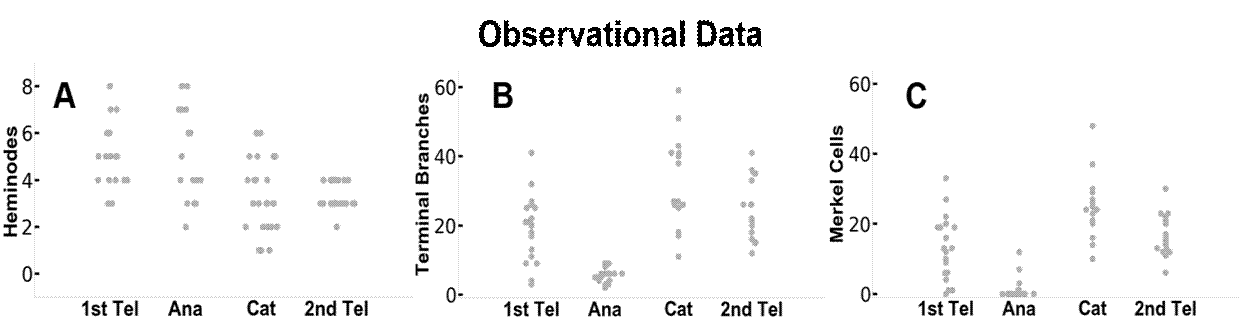
Abbildung 3. Vergleich zwischen den tatsächlichen Beobachtungsdaten (A-C) und den Daten des DES-Modells (D-F). Abgebildet sind Daten zu den einzelnen Heminodien, terminalen Neuritenverzweigungen und Merkelzellen in jedem der vier Stadien des Haarzyklus. In den Feldern D-F werden blaue bzw. orangefarbene Punkte verwendet, um zwei getrennte Endorgane durch ihre Stadien des Haarzyklus zu verfolgen. Dieser Stil ist auch mit den Daten in Abbildung 4 verbunden.
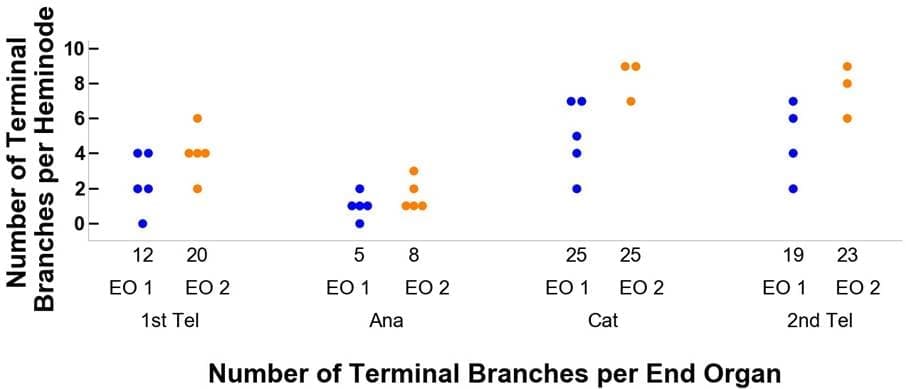
Die Endorgane, die durch die orangefarbenen und blauen Punkte in den Feldern D-F von Abbildung 3 gekennzeichnet sind, werden in Abbildung 4 näher erläutert. In Abbildung 4A beispielsweise beginnt Endorgan 1 (EO 1) im ersten Telogen mit zwölf Endverzweigungen und fünf Heminknoten, wobei die Endverzweigungen auf die Heminknoten mit 4, 4, 2, 2, 0 pro Heminknoten verteilt sind.Wenn Endorgan 1 dann in die anagene Phase des Haarzyklus übergeht, verringert sich die Anzahl der terminalen Äste auf fünf. Diese fünf terminalen Äste sind auf die fünf Heminknoten mit einer Entsprechung von 2, 1, 1, 1, 0 verteilt. Außerdem ist zu beachten, dass beim Übergang vom Katagen zum zweiten Telogen die Anzahl der Heminknoten für dieses Endorgan von fünf auf vier sinkt.
Es wurde eine Sensitivitätsanalyse durchgeführt, um herauszufinden, welche Regeln einen größeren Einfluss auf das Umgestaltungsverhalten der Endorgane haben. Zunächst wurden die Regeln untersucht, die mit der terminalen Verzweigung und der Entfernung der Merkelzellen vom ersten Telogen zum Anagen verbunden sind. Während des Übergangs hängt die Löschung eines terminalen Astes davon ab, ob der terminale Ast mit einer Merkelzelle besiedelt ist oder nicht. Bei nicht besiedelten Endverzweigungen führte eine Erhöhung der Löschungswahrscheinlichkeit um 0,1 zu einem Unterschied von 20,5 % in der Gesamtzahl der Endverzweigungen, während die gleiche Änderung bei besiedelten Endverzweigungen zu einem Unterschied von 1 % führte. Außerdem wurden zwei Regeln für die Beschneidung der Terminusäste vom Katagen zum zweiten Telogen bewertet. Die Analyse ergab, dass sich die Anzahl der Heminodien im Durchschnitt um 6,7 % ändert, wenn die Wahrscheinlichkeit einer Heminodendeletion um 0,05 für Heminodien ohne Merkelzellen steigt. Andererseits führte die gleiche Änderung bei besiedelten Heminodien nur zu einem durchschnittlichen Unterschied von 1,6 % in der Anzahl der Heminodien. Aus diesen Ergebnissen lässt sich schließen, dass die Anzahl der Endorgan-Bestandteile empfindlicher auf die Löschungsmaßnahmen von Endorgan-Bestandteilen reagiert, die weniger besiedelt sind.
(A)
(B)
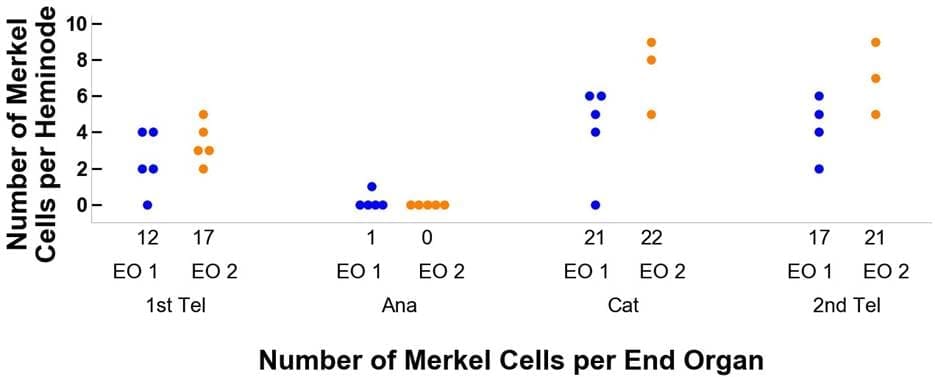
Abbildung 4. Verfolgung des Verlaufs von zwei Beispielendorganen durch ihre Stadien des Haarzyklus vom ersten Telogen, Anagen und Katagen bis zum zweiten Telogen. Tafel A zeigt die Veränderungen in den Endverzweigungen und Tafel B bezieht sich auf die Merkelzellen. Die blaue Farbe steht für Endorgan 1 (EO 1) und orange (EO 2). Endorgan 1 beginnt beispielsweise mit zwölf Endverzweigungen und zwölf Merkelzellen.Beim Übergang zum Anagenstadium des Haarzyklus sinkt die Anzahl der Endverzweigungen auf 5 und die der Merkelzellen auf 1, mit einer Verteilung auf fünf Heminknoten von 2, 1, 1, 1, 1, 0 bzw. 1, 0, 0, 0, 0.
Diskussion
Wie in der Einleitung erwähnt, wurde in dieser Arbeit versucht, mit Hilfe eines DES-Ansatzes die Prinzipien zu ermitteln, die bestimmen, wie sich die Äste im Verlauf des Haarzyklus verändern, und die Strategien der Merkelzellen- und Heminodendynamik zu bewerten. Die Wahl eines Bottom-up-Ansatzes, bei dem Endorgane, Heminoden und terminale Neuritenäste als unterschiedliche Entitäten modelliert werden, ermöglichte es uns, die herrschenden Strategien zu verfeinern und optimale Parameter abzuschätzen, die bei den Umbauprozessen eine Rolle spielen. Das Modell scheint die beobachteten Daten gut zu replizieren und auf dieser Abstraktionsebene über eine Reihe von Wahrscheinlichkeitswerten zu spekulieren, die die biologischen Prozesse steuern könnten.Durch die Simulation verschiedener Kombinationen von Geschwindigkeitsentwicklungen und Inkorporationsregeln konnten wir die Dynamik in den Endorganarmen darstellen, um sie mit spontanen Haarzyklusbeobachtungen zu vergleichen und die Lücken zu füllen, die durch die Beschränkungen in Beobachtungsstudien entstanden sind. Beispielsweise ermöglichte der entitätsbasierte Bottom-up-Ansatz eine detaillierte Rückverfolgbarkeit der Endorgane und ihrer einzelnen Komponenten durch jede Phase des Haarzyklus, was in der Beobachtungsforschung nur eingeschränkt möglich ist. Außerdem konnten wir mit Hilfe des DES-Modells für jeden Bestandteil des Endorgans über alle Stadien des Haarzyklus hinweg verfeinerte Wahrscheinlichkeiten für die Hinzufügung und Löschung bestimmen. Allerdings wurden beim Aufbau des DES-Modells mehrere Annahmen getroffen. Erstens wurde davon ausgegangen, dass die beobachteten Mittelwerte im Datensatz über den spontanen Haarzyklus der Hintergliedmaßen genau genug sind, um logische Strategien zu entwickeln. Zweitens wurde davon ausgegangen, dass Heminodien mit der geringsten Anzahl von Merkel-Zellen am ehesten entfernt werden. Andererseits wurde angenommen, dass Merkel-Zellen in großen Merkel-Zell-Clustern am ehesten entfernt werden. Schließlich wurde angenommen, dass Merkel-Zellen zufällig zu Heminknoten hinzugefügt werden.
Die Anwendung eines DES-Ansatzes auf Probleme in der biologischen Forschung ist im Vergleich zu seinen Anwendungen in anderen Bereichen (Hunt 2009), wie z. B. im Gesundheitswesen im weiteren Sinne, begrenzt gewesen. Diese Studie zeigt, dass DES eine vielversprechende Ergänzung zu den gängigen Forschungspraktiken in der biologischen Forschung sein kann. Indem man potenzielle biologische Regeln testet, bevor man zu Mausmodellen übergeht, können sowohl Zeit als auch Geld gut eingesetzt werden. Verschiedene Was-wäre-wenn-Szenarien und Sensitivitätsanalysen unter Verwendung von DES-Modellen können auch dazu beitragen, zusätzliche Experimente zu entwerfen, indem sie über kritische Faktoren informieren, die eng mit der emergenten Dynamik zusammenhängen. Dieses Modell soll beispielsweise die Möglichkeit bieten, mit Parametern zu experimentieren. Der Anreiz, der hinter einem solchen Modell steht, ist die Untersuchung möglicher verwirrender experimenteller Faktoren, wie z. B. der Zeitpunkt und die Bedingungen des Endorgans, die das Verhalten eines Endorgans und das Verhalten seiner Bestandteile während des Haarzyklus potenziell beeinflussen könnten. Das Experimentieren kann insbesondere durch die Simulation dieser Unregelmäßigkeiten in Kombination mit den Wahrscheinlichkeitsregeln des Basismodells erfolgen, um die tatsächliche Robustheit der ermittelten probabilistischen Regeln zu bestimmen.
Biographien der Autoren
HYOJUNG KANGist wissenschaftliche Assistenzprofessorin in der Abteilung für System- und Informationstechnik an der Universität von Virginia. Sie hat in Wirtschaftsingenieurwesen und Operations Research an der Penn State University promoviert. Zu ihren Forschungsinteressen gehören die Entwicklung und Anwendung hybrider Simulationsmodelle, um umsetzbare Lösungen für komplexe Systeme, insbesondere im Gesundheitswesen, zu finden. Ihre E-Mail Adresse lautet hkang@virginia.edu.
RACHEL L. ORLOWSKY ist Studentin in den Abteilungen für System- und Informationstechnik und Biomedizinische Technik an der University of Virginia. Ihr Interesse gilt der computergestützten Modellierung in biologischen Bereichen. Ihre E-Mail-Adresse lautet rlo4kc@virginia.edu.
GREGORY J. GERLING ist außerordentlicher Professor in den Abteilungen für System- und Informationstechnik und Biomedizinische Technik an der Universität von Virginia. Er promovierte an der University of Iowa und beschäftigt sich in seiner Forschung mit Computational Neuroscience, Haptik, Human Factors und Mensch-Maschine-Interaktion. Sein Hauptinteresse gilt dem Verständnis der Komplexität des taktilen Sinnessystems, das unsere Wahrnehmung von Berührungen und die Entwicklung von haptischen Geräten beeinflusst. Seine E-Mail Adresse lautet gg7h@virginia.edu.