El desafío
por Hyojung Kang, Rachel Orlowsky y Gregory J. Gerling (Universidad de Virginia)
Tal y como se presentó en la Conferencia de Simulación de Invierno de 2017
En los mamíferos, el tacto está codificado por receptores sensoriales incrustados en la piel. Para una clase de receptores en el ratón, la arquitectura de sus células de Merkel, neuritas no mielinizadas y heminodos siguen tendencias particulares de renovación y remodelación a lo largo de las etapas del ciclo del pelo desde las 4 a las 10 semanas de edad. Como actualmente es imposible observar tales tendencias a lo largo del ciclo del pelo de un solo animal, este trabajo emplea la simulación de eventos discretos para identificar y evaluar las políticas de la dinámica de las células de Merkel y los heminodos. Los resultados, que se ajustan bien a los datos observados, muestran que el modelo de referencia reproduce comportamientos de remodelación dinámica entre las fases del ciclo piloso, basándose en políticas particulares de adición y eliminación y en probabilidades estimadas vinculadas a las partes constituyentes de las células de Merkel, las neuritas de las ramas terminales y los heminodos. El análisis muestra además que ciertas políticas tienen mayor influencia que otras. Este uso de la computación es un enfoque novedoso para comprender el desarrollo neuronal.
Introducción
El sentido del tacto es clave para comportamientos de la vida cotidiana como alimentarse, establecer vínculos sociales y evitar daños corporales. En los mamíferos, el tacto está codificado por receptores sensoriales incrustados en la piel (Kandel 2012). Los receptores sensoriales incluyen aferentes cutáneos de tacto ligero, así como los que señalan información relativa a la propiocepción, quimiorrecepción y dolor. Tanto los receptores sensoriales como la piel se renuevan y remodelan continuamente para mantener la barrera en estados normales y tras una lesión (Chung 2010; Marshall 2016; Müller-Röver 2001; Rajan 2003). En la piel vellosa, las terminaciones nerviosas de las células de Merkel se agrupan en estructuras epiteliales especializadas denominadas "cúpulas táctiles" (Plikus 2008). Los ratones tienen cientos de cúpulas táctiles en su piel vellosa y los humanos también tienen terminaciones nerviosas similares, aunque sutilmente diferentes. En términos más generales, los receptores de las células de Merkel se encuentran tanto en la piel vellosa como en la glabra de los mamíferos, aunque las estructuras locales de los receptores y de la piel varían en cada caso. En las poblaciones receptivas, estos aferentes ayudan a señalar la información relativa a los bordes y la curvatura de los estímulos, entre otros atributos (Johnson 2001).
La dinámica de la arquitectura del complejo célula de Merkel-neurita apenas está empezando a comprenderse (obsérvese la abstracción dada en la Figura 1, y Lesniak 2014). Se ha observado que la conexión de una célula de Merkel con una neurita terminal o su separación de ella, así como las neuritas de los heminodos, siguen tendencias específicas de las fases del ciclo piloso en el ratón (Marshall 2016). En particular, a medida que los ratones envejecen, se observan múltiples ciclos pilosos sincronizados, en los que el pelo del animal cambia en todo su cuerpo de forma ondulada (Müller-Röver 2001). Existen cuatro etapas en el ciclo espontáneo del pelo de los ratones: Primera Telógena: 4 semanas, Anágena: 5-6 semanas, Catágena: 6 semanas, y Segunda Telógena: 9-10 semanas. Después de este punto, el ciclo del pelo empieza a entrar en una fase de mosaico en la que el pelo del cuerpo del animal no cambia de forma ondulada, sino que se pierde y vuelve a crecer a diferentes velocidades en posiciones aparentemente aleatorias de la superficie de la piel. Las neuronas cutáneas y las células de Merkel pueden participar en mecanismos de plasticidad durante la regeneración de los folículos pilosos; sin embargo, la dinámica y las consecuencias fisiológicas de la plasticidad neuronal en los receptores táctiles no se conocen del todo (Moll 1996; Nakafusa 2006).
Los esfuerzos de investigación observacional en relación con la remodelación de las ramas son limitados, ya que actualmente no podemos rastrear órganos terminales específicos a lo largo del ciclo del pelo. Un enfoque de modelado, por el contrario, puede permitir un seguimiento detallado de los órganos terminales y cada uno de sus componentes a través de cada etapa del ciclo del pelo. Por otra parte, a pesar de los casos de modelos de simulación de eventos discretos (DES) en la investigación biológica, todavía hay muy pocos ejemplos publicados de modelos validados exhaustivamente. Por ello, nuestro grupo construyó recientemente un modelo computacional para probar un conjunto inicial de reglas que rigen los mecanismos de remodelación de los arbores (Marshall 2016). Con su enfoque descendente, se utilizaron las estadísticas de población de los datos observados para construir el modelo computacional. Utilizando las estadísticas de población como puntos de referencia, se crearon y eliminaron iterativamente constituyentes de órganos finales para reflejar los datos observados. Cada una de las transiciones entre las fases del ciclo piloso se modeló como una simulación independiente. Dentro de cada simulación, había una serie de iteraciones en las que las células de Merkel y/o los heminodos podían añadirse o eliminarse de acuerdo con sólo cuatro políticas probabilísticas, informadas por los datos morfométricos. Ese esfuerzo identificó cuatro políticas de dinámica de células de Merkel y heminodes a partir de datos observacionales y evaluó las políticas.
Aunque el estudio (Marshall 2016) contribuyó a explicar las reglas que rigen los procesos de remodelación de las neuritas (o arboretes) de las células de Merkel, esos modelos computacionales tienen varias limitaciones. Por ejemplo, los constituyentes individuales de los órganos terminales de las células de Merkel, las neuritas terminales y los heminodos no eran rastreables entre las fases del ciclo piloso. Además, el hecho de estar limitados por un enfoque descendente dificultaba la formación de políticas de remodelación de los arborescentes centradas en los componentes individuales del órgano terminal, en lugar de en el propio órgano terminal. Asimismo, debido a la estructura del modelo, no era fácil realizar pruebas de escenarios hipotéticos y determinar los parámetros óptimos asociados a las reglas de gobierno. Por último, este modelo carece de funciones sencillas de optimización o experimentación, lo que restringe el ensayo de nuevos parámetros no definidos previamente. Por lo tanto, hemos intentado superar estas limitaciones empleando un enfoque DES. En concreto, el objetivo de este estudio es demostrar cómo un enfoque de modelado DES puede ayudar a comprender los mecanismos subyacentes de los cambios en las ramas a lo largo del ciclo piloso. En contraste con los esfuerzos previos de modelado computacional, el que aquí se presenta adopta un enfoque ascendente, con reglas de probabilidad codificadas para cada constituyente específico a medida que el órgano terminal progenitor atraviesa el ciclo piloso. Las políticas son una combinación de reglas condicionales y probabilísticas. A diferencia del modelo computacional, cada célula de Merkel, rama neurítica terminal y heminodo relacionado con un órgano terminal concreto se examinó individualmente, y las reglas que rigen su adición y supresión reflejaban hipótesis relativas a las características biológicas. Este enfoque ascendente, que modela los órganos terminales, los heminodos y las ramas neuríticas terminales como entidades diferentes, nos permitió perfeccionar las políticas rectoras y estimar los parámetros óptimos que intervienen en los procesos de remodelación. Las células de Merkel se modelan como un atributo de las ramas terminales que afecta a las políticas y los parámetros.
La solución
Método
El objetivo de este trabajo es utilizar el DES para identificar los principios que especifican cómo cambian las ramas a lo largo del ciclo piloso y evaluar las políticas de la dinámica de las células de Merkel y los heminodos. Se simulan diferentes combinaciones de evoluciones de velocidad y reglas de incorporación para obtener predicciones de las ramas de los órganos terminales y compararlas con las observaciones espontáneas del ciclo piloso.
Abstracción de la biología para su uso en la descripción de las entradas, salidas y transiciones modeladas
Si bien la arquitectura del complejo célula de Merkel-neurita puede ser aún más complicada en términos de sus partes constituyentes, el nivel de abstracción dado en la Figura 1 es algo típico y se utilizará para describir las entradas y salidas del modelo que se construirá en este documento.

Sobre la base del esfuerzo de modelado anterior (Marshall 2016), se desarrolló un conjunto preliminar de reglas, que se han modificado en el presente trabajo para venir de una perspectiva de abajo hacia arriba, como se muestra en la Tabla 1. En general, comprobamos primero el estado de un heminodo comprobando las ramas terminales asociadas y las células de Merkel.
Simulación de eventos discretos
Descripción del modelo
Se construyó un modelo DES para reproducir la dinámica que se produce durante las transiciones del ciclo capilar y determinar los parámetros clave que afectan a los comportamientos dinámicos. El modelo constaba de tres tipos de entidades, cada una de las cuales representaba órganos terminales, heminodos y ramas terminales. Las células de Merkel se modelaron como un atributo binario de las entidades de las ramas terminales: cada rama terminal o bien tiene una célula de Merkel (1) o bien no la tiene (0). La estructura de las ramas se modeló como una jerarquía de dos capas de lotes. Cada entidad de órgano terminal consta de un lote de entidades de heminodo, y cada entidad de heminodo consta de un lote de entidades de rama terminal. A medida que cada órgano terminal atraviesa un ciclo capilar completo, su composición cambia continuamente mediante procesos de dosificación/desdosificación con diferentes reglas. El modelo se construyó utilizando el paquete de software de simulación y programación Simio.
Estimación y validación de parámetros
Para desarrollar el modelo de referencia, utilizamos datos observacionales obtenidos del estudio anterior (Marshall 2016). Los principales parámetros de First Telogen incluían el número de heminodes por órgano terminal, el número de ramas terminales por órgano terminal y el número de células de Merkel por órgano terminal, el número de ramas terminales por heminode y el número de células de Merkel por heminode. Estos parámetros se estimaron utilizando distribuciones empíricas. La duración de cada proceso fue de 4, 2 y 4 semanas entre las etapas de los ciclos pilosos, respectivamente.
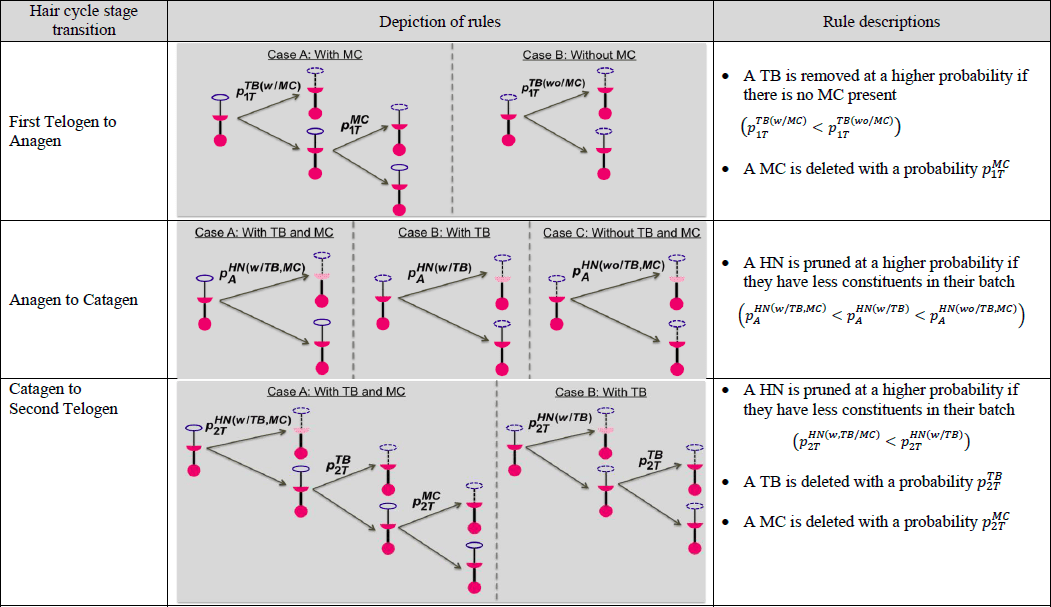
Tabla 1. Representación gráfica y textual de las reglas asociadas a cada una de las tres transiciones del ciclo piloso, donde MC = célula de Merkel y TB = rama terminal y donde![]() representa una probabilidad de que un componente de un órgano final xsea eliminado/ podado en la etapas xincluye un MC, TB y HN. El términos incluye 1T (Primer Telógeno), A (Anágeno), C (Catágeno) y 2T (Segundo Telógeno).
representa una probabilidad de que un componente de un órgano final xsea eliminado/ podado en la etapas xincluye un MC, TB y HN. El términos incluye 1T (Primer Telógeno), A (Anágeno), C (Catágeno) y 2T (Segundo Telógeno).
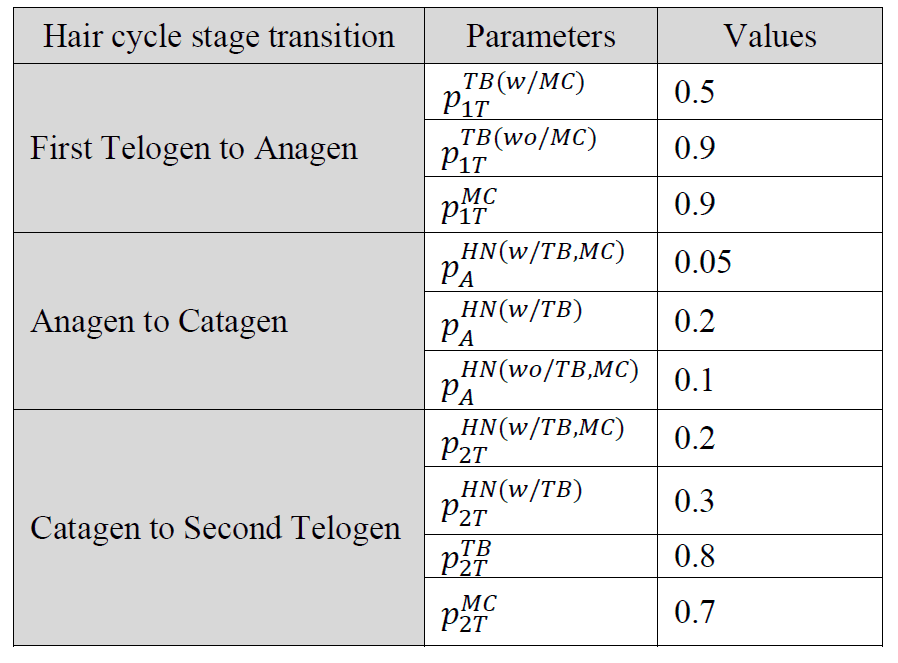
Uno de los objetivos de este estudio era estimar las probabilidades de adición y supresión de cada célula de Merkel, rama terminal y heminodio relativo a un órgano terminal entre etapas. Para calibrar esos parámetros desconocidos del modelo, se consideraron tres resultados principales: el número medio de heminodes por órgano terminal, el número medio de células de Merkel por órgano terminal y el número medio de células de Merkel por heminode. La calibración del modelo se realizó en cada estadio cronológicamente, de uno en uno, porque las probabilidades en los distintos estadios son independientes entre sí, pero las estadísticas de población de cada estadio del ciclo piloso dependen del estadio anterior. El modelo de referencia calibrado se ejecutó con 25 réplicas y se validó comparando sus resultados con los tres resultados principales de los datos observacionales. Se realizó un análisis de sensibilidad para comprender el impacto de los cambios en las probabilidades de supresión/adición en los principales resultados entre el catágeno y el segundo telógeno. Elegimos las dos etapas porque durante la última transición del ciclo piloso se producen más actividades de remodelación que afectan a los heminodos, las ramas terminales y las células de Merkel, en comparación con otras transiciones.
El impacto comercial
Resultados
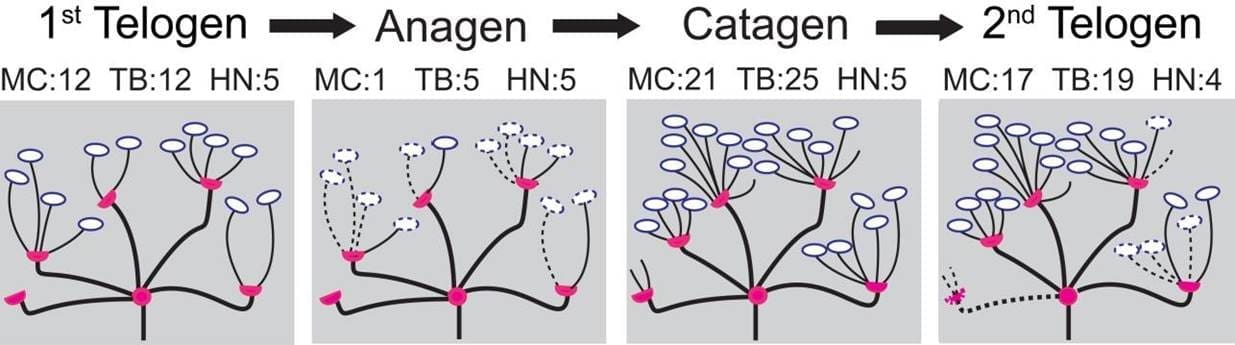
En la figura 2 se muestra un ejemplo de simulación de un órgano terminal a través de las cuatro etapas del ciclo del cabello. Basándonos en las políticas conceptuales de eliminación/adición entre ciclos capilares desarrolladas en el estudio anterior (Marshall 2016), hemos generado probabilidades ascendentes como se muestra en la Tabla 2 (página siguiente). En la transición de Primer Telógeno a Anágeno, la probabilidad de que se elimine una rama terminal es de 0,9 si no contiene una célula de Merkel y la probabilidad es de 0,5 si hay una célula de Merkel. Durante la transición, se elimina una célula de Merkel con una probabilidad de 0,9. En la transición de Anágena a Catágena, los heminodos tienen más probabilidades de ser podados si sus constituyentes están vacíos. Por ejemplo, un heminodo se poda con una probabilidad de 0,2 si no contiene ninguna rama terminal células de Merkel, con una probabilidad de 0,1 si tiene ramas terminales pero no células de Merkel, y con una probabilidad de 0,05 si contiene tanto ramas terminales como células de Merkel. En la transición de Catágena a Segunda Telógena, los heminodos siguen una política de eliminación proporcional similar. Durante la transición, los heminodos pierden al menos una rama terminal con una probabilidad de 0,8. El número de ramas terminales que se suprimen oscila entre 1 y 3 con la misma probabilidad. Una célula Merkel se elimina con una probabilidad de 0,9.
El modelo de simulación de referencia se validó utilizando los datos observacionales recopilados por Marshall et al. (2016). En la tabla 3 se compara el número medio de células de Merkel y heminodes en cada etapa derivado de 20 datos observacionales con los estimados a partir del modelo DES con 20 réplicas. Los resultados del modelo fueron considerablemente similares a los datos reales, lo que indica que el modelo de simulación de referencia es una representación razonable del sistema arbóreo (Banks 2000).
Tabla 2. Valores de los parámetros utilizados en el modelo de simulación en cada una de las tres transiciones del ciclo piloso, donde MC = célula de Merkel y TB = rama terminal y donde![]() representa una probabilidad de que un componente de un órgano terminal xsea eliminado/ podado en la etapas xincluye un MC, TB y HN. El términos incluye 1T (Primer Telógeno), A (Anágeno), C (Catágeno) y 2T (Segundo Telógeno).
representa una probabilidad de que un componente de un órgano terminal xsea eliminado/ podado en la etapas xincluye un MC, TB y HN. El términos incluye 1T (Primer Telógeno), A (Anágeno), C (Catágeno) y 2T (Segundo Telógeno).
|
Datos observacionales (media) |
Resultados del modelo (media [IC 95 %]) |
|||
|
Células de Merkel |
Heminodes |
Células de Merkel |
Heminodes |
|
| Primer telógeno |
13 |
5 |
13 [12.4, 17.3] |
5 [5.3, 5.5] |
| Anágena |
0 |
5 |
0 [0, 0.7] |
5 [5.2, 5.5] |
| Catágena |
25 |
4 |
24 [24.1, 24.8] |
4 [3.9, 4.2] |
| Segundo telógeno |
15 |
3 |
14 [13.8, 14.2] |
3 [3.2, 3.9] |
La figura 3 compara los resultados de los datos observados y simulados. Al basarse únicamente en señales relacionadas con el ciclo del pelo, la dinámica del modelo produjo ramas que concuerdan estrechamente con las observaciones experimentales. Las distribuciones de heminodos, ramas terminales y células de Merkel por órgano terminal en cada ciclo piloso y los patrones de cambios fueron similares entre los datos observacionales y los resultados de la simulación. El modelo de simulación también nos permitió rastrear cada órgano terminal a lo largo del ciclo piloso. Por ejemplo, los puntos azules y naranjas de los paneles D, E y F de la Figura 3 representan dos órganos terminales diferentes, respectivamente. Sin embargo, los comportamientos dinámicos de remodelación entre la observación experimental y los resultados del modelo no se compararon a nivel de órgano terminal debido a la falta de trazabilidad en los datos observacionales.
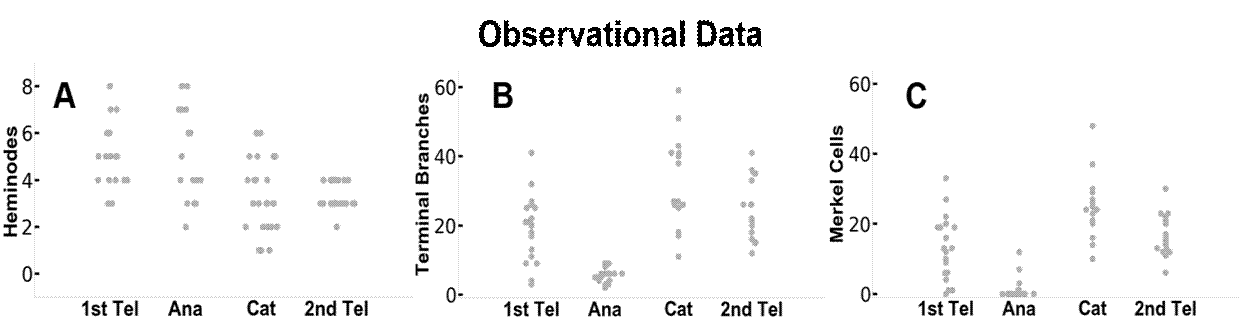
Figura 3. Comparación entre (A-C) Comparación entre los datos observacionales reales (A-C) y los datos del modelo DES (D-F). Se muestran los datos relativos a cada uno de los heminodos, las ramas terminales de las neuritas y las células de Merkel a lo largo de cada una de las cuatro etapas del ciclo piloso. En los paneles D-F, se utilizan puntos azules y naranjas, respectivamente, para trazar dos órganos finales separados a través de sus etapas del ciclo piloso. Este estilo se vincula también a los datos de la figura 4.
Los órganos finales denotados por puntos naranjas y azules marcados en los paneles D-F de la Figura 3 se detallan más en la Figura 4. Por ejemplo, en la Figura 4A, el órgano final 1 (OE 1) comienza en el Primer Telógeno con doce ramas terminales y cinco heminodos donde las ramas terminales se distribuyen a través de los heminodos con 4, 4, 2, 2, 0 por cada heminodos.Después, cuando el órgano terminal 1 pasa a la fase Anágena del ciclo piloso, su número de ramas terminales disminuye a cinco. Estas cinco ramas terminales se distribuyen por los cinco heminodos con una correspondencia de 2, 1, 1, 1, 0. Además, nótese que en la transición de Catágena a Segunda Telógena el número de heminodos disminuye de cinco a cuatro para este órgano terminal.
Se realizó un análisis de sensibilidad para comprender qué reglas tienen una mayor influencia en el comportamiento de remodelación del órgano terminal. En primer lugar, se examinaron las reglas asociadas a la eliminación de la rama terminal y de las células de Merkel del Primer Telógeno al Anágeno. Durante la transición, la eliminación de una rama terminal depende de si la rama terminal está poblada con una célula de Merkel o no. En el caso de las ramas terminales no pobladas, un aumento de 0,1 en la probabilidad de deleción dio lugar a una diferencia del 20,5% en el número total de ramas terminales, mientras que el mismo cambio para las ramas terminales pobladas dio lugar a una diferencia del 1%. También se evaluaron dos reglas que rigen la poda de heminodo desde el catágeno hasta el segundo telógeno. El análisis mostró que al aumentar en 0,05 la probabilidad de supresión de heminodos para los heminodos sin células de Merkel, el número de heminodos cambiaba en un 6,7% de media. Por otra parte, el mismo cambio para los heminodes poblados sólo dio lugar a una diferencia media del 1,6% en el número de heminodes. A partir de estos resultados, podemos concluir que la cantidad de heminodos es más sensible a las políticas de supresión de heminodos menos poblados.
(A)
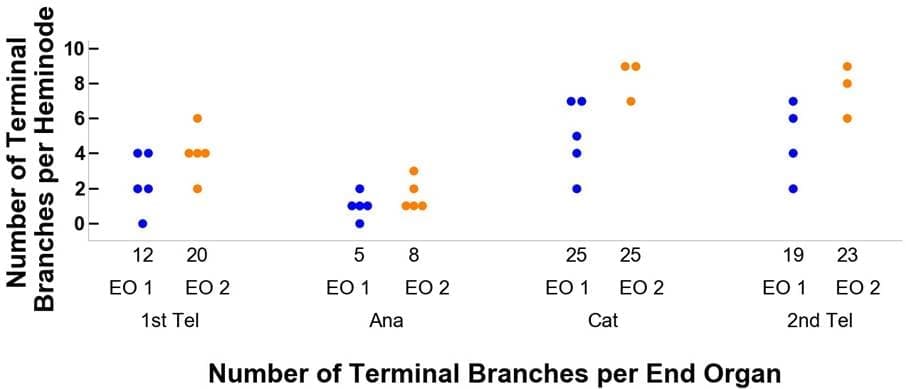
(B)
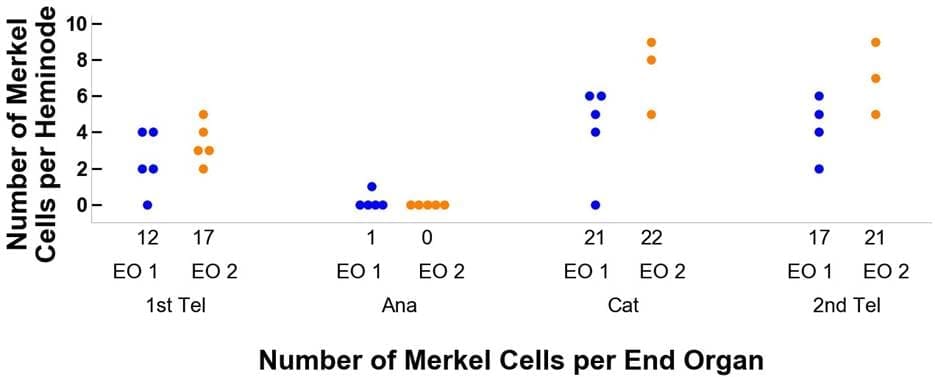
Figura 4. Trazado del flujo de dos órganos terminales de ejemplo a través de sus fases del ciclo piloso, desde la primera telógena, anágena y catágena, hasta la segunda telógena. El panel A detalla los cambios en las ramas terminales y el panel B se refiere a las células de Merkel. El color azul representa el órgano terminal 1 (OE 1) y el naranja el OE 2. Por ejemplo, el órgano terminal 1 comienza con doce ramas terminales y doce células de Merkel.A continuación, al pasar a la fase anágena del ciclo piloso, el número de ramas terminales disminuye a 5 y el de células de Merkel a 1, con una distribución en cinco heminodos de 2, 1, 1, 1, 0 y 1, 0, 0, 0, 0, respectivamente.
Discusión
Como se señaló en la introducción, este trabajo trató de emplear un enfoque DES para identificar los principios que especifican cómo arbors cambiar en el transcurso del ciclo del cabello y para evaluar las políticas de las células de Merkel y heminode dinámica. La elección de un enfoque ascendente, que modela los órganos terminales, los heminodos y las ramas terminales de las neuritas como entidades diferentes, nos permitió refinar las políticas gobernantes y estimar los parámetros óptimos que desempeñan un papel en los procesos de remodelación. El modelo parece replicar bien los datos observados y especular sobre un conjunto de valores probabilísticos que podrían estar impulsando los procesos biológicos, a este nivel de abstracción.La simulación de distintas combinaciones de evoluciones de velocidad y reglas de incorporación nos permitió representar la dinámica en las ramas de los órganos terminales para compararla con las observaciones espontáneas del ciclo piloso y ayudó a llenar las lagunas dejadas por las limitaciones de los estudios de investigación observacionales. Por ejemplo, el enfoque ascendente basado en entidades permitió un seguimiento detallado de los órganos terminales y de cada uno de sus componentes a lo largo de cada etapa del ciclo piloso, lo que está restringido en la investigación observacional. Además, gracias al modelo DES, pudimos determinar probabilidades precisas de adición y supresión de cada componente del órgano final en cada una de las fases del ciclo piloso. Dicho esto, se hicieron varias suposiciones al construir el modelo DES. En primer lugar, se asumió que los valores medios observados en el conjunto de datos del ciclo piloso espontáneo de la extremidad posterior son lo suficientemente precisos como para elaborar políticas lógicas. En segundo lugar, se supuso que los heminodos con el menor número de células de Merkel tenían más probabilidades de ser eliminados y, por otro lado, que las células de Merkel en grandes grupos de células de Merkel tenían más probabilidades de ser eliminadas. Por último, se supuso que las células de Merkel se añaden aleatoriamente a los heminodos.
Las aplicaciones de un enfoque DES a problemas de investigación biológica han sido limitadas en comparación con sus aplicaciones en otras áreas (Hunt 2009), como la atención sanitaria en general. Este estudio demuestra que el DES puede utilizarse como un complemento prometedor a las prácticas de investigación comúnmente utilizadas en la investigación biológica. Al probar las posibles reglas biológicas antes de pasar a los modelos de ratón, tanto el tiempo como el dinero podrían emplearse bien. Diversas pruebas de escenarios hipotéticos y análisis de sensibilidad utilizando modelos DES también pueden ayudar a diseñar experimentos adicionales informando sobre factores críticos estrechamente relacionados con la dinámica emergente. Por ejemplo, este modelo pretende ofrecer características de experimentación de parámetros. El incentivo de este modelo es estudiar posibles factores experimentales de confusión, como el momento y las condiciones del órgano final, que podrían influir en el comportamiento de un órgano final y en el de sus componentes a lo largo del ciclo capilar. Específicamente, la experimentación puede transpirar a través de la simulación de esas irregularidades en combinación con las reglas de probabilidad del modelo base para determinar la robustez real de las reglas probabilísticas determinadas.
Biografías de los autores
HYOJUNG KANGes profesora ayudante de investigación en el Departamento de Ingeniería de Sistemas e Información de la Universidad de Virginia. Es doctora en Ingeniería Industrial e Investigación Operativa por la Universidad Estatal de Pensilvania. Sus intereses de investigación incluyen el desarrollo y la aplicación de modelos de simulación híbridos para proporcionar soluciones implementables para sistemas complejos, en particular los sistemas sanitarios. Su dirección de correo electrónico es hkang@virginia.edu.
RACHEL L. ORLOWSKY es estudiante universitaria en los Departamentos de Ingeniería de Sistemas e Información y de Ingeniería Biomédica de la Universidad de Virginia. Sus intereses se centran en la modelización computacional aplicada a ámbitos biológicos. Su dirección de correo electrónico es rlo4kc@virginia.edu.
GREGORY J. GERLING es Profesor Asociado de los Departamentos de Ingeniería de Sistemas e Información y de Ingeniería Biomédica de la Universidad de Virginia. Obtuvo su doctorado en la Universidad de Iowa. Sus intereses de investigación incluyen la neurociencia computacional, la háptica, los factores humanos y la interacción hombre-máquina. Su principal interés radica en comprender la complejidad que subyace en el sistema sensorial táctil, que informa nuestra percepción del tacto y el diseño de dispositivos hápticos. Su dirección de correo electrónico es gg7h@virginia.edu.