挑战
作者:Hyojung Kang、Rachel Orlowsky 和 Gregory J. Gerling(弗吉尼亚大学)
在 2017 年冬季模拟大会上发表
在哺乳动物中,触觉是由嵌入皮肤中的感觉受体编码的。 对于小鼠的一类受体,其梅克尔细胞、无髓鞘神经元和heminode的结构在4至10周龄的毛发周期阶段中遵循特定的更新和重塑趋势。 由于目前不可能在单个动物的毛发周期中观察到这种趋势,这项工作采用离散事件模拟来识别和评估梅克尔细胞和heminode动态的政策。结果表明,基线模型根据特定的添加和移除策略,以及与梅克尔细胞、末端分支神经元和heminode的构成部分相关的估计概率,复制了毛发周期各阶段之间的动态重塑行为。 分析进一步表明,某些策略比其他策略具有更大的影响力。 这种计算方法的使用是了解神经元发育的一种新方法。
导言
触觉是日常生活行为的关键,如进食、社会联系和避免身体伤害。在哺乳动物中,触觉由嵌入皮肤的感觉受体编码(Kandel,2012 年)。感觉受体包括皮肤轻触传入以及有关本体感觉、化学感觉和疼痛的信号信息。感觉受体和皮肤都在不断更新和重塑,以维持正常状态和受伤后的屏障(Chung,2010 年;Marshall,2016 年;Müller-Röver,2001 年;Rajan,2003 年)。在多毛皮肤中,梅克尔细胞神经末梢聚集在被称为 "触摸穹顶 "的特化上皮结构中(Plikus,2008 年)。小鼠的多毛皮肤上有数以百计的触摸穹顶,人类也有类似的神经末梢,但又有细微差别。 从更广泛的意义上讲,梅克尔细胞受体遍布哺乳动物的多毛和无毛皮肤,但每种情况下的局部受体和皮肤结构各不相同。 在受体群中,此类传入有助于发出有关刺激物边缘和弧度等属性的信号(Johnson,2001 年)。
人们刚刚开始了解梅克尔细胞-神经元复合体的动态结构(注意图 1 中的抽象概念,以及 Lesniak 2014)。据观察,梅克尔细胞与末端神经元的连接或从末端神经元中移除,以及来自heminodes的神经元,都遵循小鼠毛发周期各阶段的特定趋势(Marshall,2016年)。特别是,随着小鼠年龄的增长,会观察到多个同步毛发周期,即动物全身的毛发以波浪状的方式发生变化(Müller-Röver,2001 年)。小鼠自发毛发周期分为四个阶段:第一脱发期:4 周;第二脱发期:5-6 周;第三脱发期:6 周;第四脱发期:9-10 周。此后,毛发周期开始进入镶嵌阶段,动物全身的毛发不再以波浪状方式变化,而是以不同的速度从皮肤表面看似随机的位置脱落和再生。皮肤神经元和梅克尔细胞可能会在毛囊再生过程中参与可塑性机制;然而,人们对触觉感受器神经元可塑性的动态和生理后果还不完全了解(Moll,1996 年;Nakafusa,2006 年)。
由于我们目前无法通过毛发周期追踪特定的末端器官,因此有关树轴重塑的观察研究工作受到了限制。与此相反,建模方法可以详细追踪毛发周期每个阶段的末端器官及其每个组成部分。此外,尽管离散事件模拟(DES)模型在生物研究中得到了广泛应用,但已发表的经过全面验证的模型实例仍然很少。因此,我们的研究小组最近建立了一个计算模型,以测试一套初步的枝轴重塑机制规则(Marshall,2016 年)。我们采用自上而下的方法,利用观测数据中的种群统计数据来构建计算模型。以群体统计数据为参考点,反复创建和删除末端器官成分,以反映观察到的数据。毛发周期阶段之间的每个过渡阶段都作为一个单独的模拟进行建模。在每次模拟中,都会进行多次迭代,根据形态测量数据,按照四种概率策略添加或删除梅克尔细胞和/或heminodes。这项工作从观测数据中确定了梅克尔细胞和heminode动态的四种策略,并对这些策略进行了评估。
虽然这项研究(Marshall,2016 年)有助于解释支配梅克尔细胞-神经节(或轴突)重塑过程的规则,但这些计算模型存在一些局限性。例如,梅克尔细胞、末端神经元和heminodes的各个末端器官成分无法在毛发周期的各个阶段之间进行追踪。此外,由于受到自上而下方法的限制,形成以末梢器官单个成分而非末梢器官本身为中心的心轴重塑策略也不容易。 而且,由于模型结构的原因,进行假设情景测试和确定与管理规则相关的最佳参数也不容易。最后,该模型没有简单的优化或实验功能,限制了对之前未定义的新参数的测试。因此,我们试图采用 DES 方法来克服这些限制。具体来说,本研究的目的是展示 DES 建模方法如何帮助理解毛发周期中树干变化的潜在机制。与之前的计算建模工作不同,本文介绍的建模工作采用自下而上的方法,在母体末端器官经历毛发周期的过程中,对每个特定成分的概率规则进行编码。策略是条件规则和概率规则的结合。与计算模型不同的是,本文对与特定末端器官相关的每个梅克尔细胞、末端神经元分支和heminode都进行了单独研究,其添加和删除规则反映了有关生物特征的假设。这种自下而上的方法将末端器官、神经节和末端神经分支作为不同的实体建模,使我们能够完善管理策略,并估算出在重塑过程中发挥作用的最佳参数。梅克尔细胞被建模为末端分支的一个属性,它会影响策略和参数。
解决方案
方法
这项工作的目的是利用 DES 来确定在毛发周期中树干如何变化的原则,并评估梅克尔细胞和heminode 的动态策略。通过模拟不同的速率演化和合并规则组合,得出末端器官轴突的预测结果,并与自发毛发周期观察结果进行比较。
生物抽象用于描述模型的输入、输出和转换
我们考虑的是小鼠多毛皮肤中的梅克尔细胞-神经元复合体。 虽然梅克尔细胞-神经元复合体的结构就其组成部分而言可能更加复杂,但图 1 所示的抽象水平具有一定的典型性,将用于描述本文所建模型的输入和输出。

一般来说,我们首先通过检查相关的末端分支和梅克尔细胞来检查heminode的状态。
离散事件模拟
模型描述
我们建立了一个 DES 模型,以再现毛发周期转换过程中发生的动态变化,并确定影响动态行为的关键参数。该模型包括三种实体类型,分别代表末端器官、heminodes 和末端分支。梅克尔细胞被模拟为末端分支实体的二元属性:每个末端分支要么有梅克尔细胞(1),要么没有梅克尔细胞(0)。轴突的结构被模拟为由两个批处理层组成的层次结构。每个末端器官实体由一批heminode实体组成,每个heminode实体由一批末端分支实体组成。该模型使用 Simio 仿真和调度软件包建立。
参数估计和验证
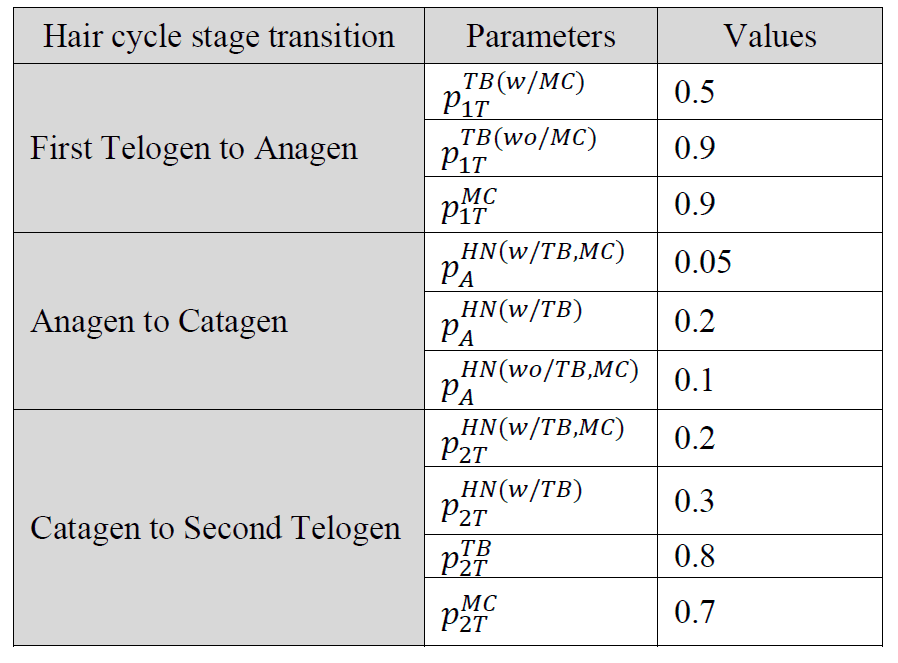
为了开发基线模型,我们使用了之前研究(Marshall,2016 年)中获得的观测数据。First Telogen 的主要参数包括:每个末端器官的heminodes数量、每个末端器官的末端分支数量,以及每个末端器官的Merkel细胞数量、每个heminode的末端分支数量和每个heminode的Merkel细胞数量。每个毛发周期阶段之间的持续时间分别为 4 周、2 周和 4 周。
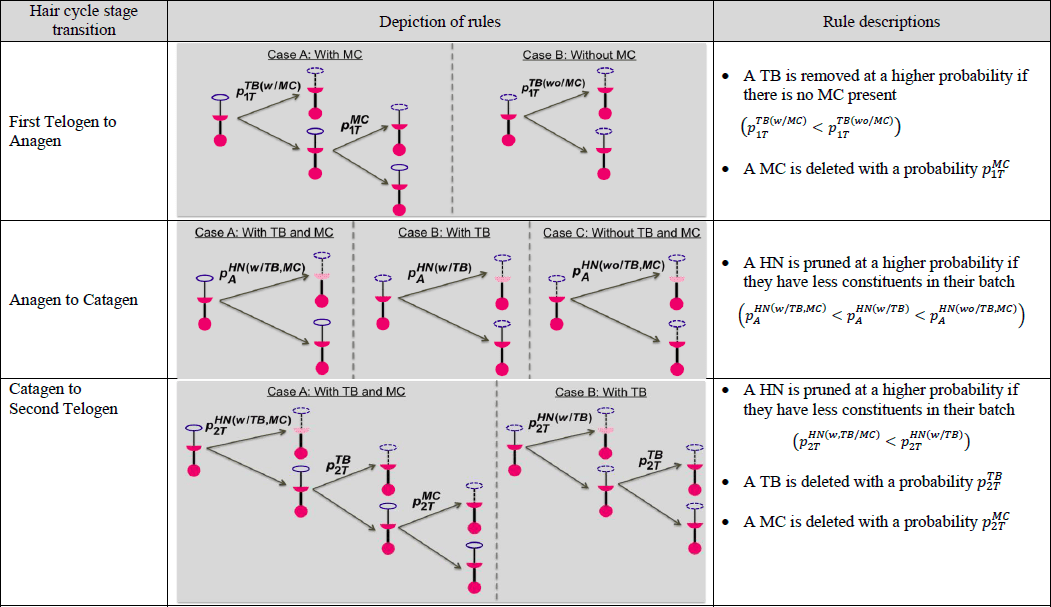
表 1.用图形和文字描述与毛发周期三个过渡阶段中每个阶段相关的规则,其中 MC = 梅克尔细胞,TB = 末端分支,![]() 代表末端器官 x 在阶段s 中被移除/修剪的概率 x包括 MC、TB 和 HN。s 阶段包括 1T(第一次脱发)、A(生长期)、C(衰老期)和 2T(第二次脱发)。
代表末端器官 x 在阶段s 中被移除/修剪的概率 x包括 MC、TB 和 HN。s 阶段包括 1T(第一次脱发)、A(生长期)、C(衰老期)和 2T(第二次脱发)。
本研究的目标之一是估算各阶段之间与终末器官相关的每个梅克尔细胞、终末分支和heminode的增加和删除概率。为了校准这些未知的模型参数,主要考虑了三个结果:每个终末器官的平均heminodes数量、每个终末器官的平均Merkel细胞数量和每个heminode的平均Merkel细胞数量。模型校准在每个阶段按时间顺序逐一进行,因为不同阶段的概率是相互独立的,但每个毛发周期阶段的种群统计都取决于前一个阶段。校准后的基线模型运行了 25 次,并通过将其结果与观测数据的三个主要结果进行比较进行了验证。我们进行了敏感性分析,以了解删除/添加概率的变化对头发生长期(Catagen)到第二衰老期(Second Telogen)之间主要结果的影响。我们之所以选择这两个阶段,是因为与其他过渡阶段相比,在毛发周期的最后一个过渡阶段会发生更多涉及heminodes、末端分支和Merkel细胞的重塑活动。
商业影响
结果
图 2 显示了一个末端器官经历毛发周期四个阶段的模拟示例。根据之前研究(马歇尔,2016 年)中制定的毛发周期之间的概念性删除/添加政策,我们生成了自下而上的概率,如表 2(下一页)所示。在从初发期向生长期过渡时,如果末端分支不包含梅克尔细胞,则该分支被删除的几率为 0.9;如果包含梅克尔细胞,则该分支被删除的几率为 0.5。在过渡期,梅克尔细胞被删除的概率为 0.9。在从 "生长期"(Anagen)到 "衰老期"(Catagen)的转变过程中,如果半节点(Heminode)的组成成分是空的,则其被修剪的可能性更大。例如,如果一个半结节不包含任何末端分支梅克尔细胞,则其被修剪的概率为 0.2;如果一个半结节包含末端分支但不包含梅克尔细胞,则其被修剪的概率为 0.1;如果一个半结节同时包含末端分支和梅克尔细胞,则其被修剪的概率为 0.05。在从衰老期向第二衰老期过渡的过程中,正常节遵循类似的比例删除策略。在过渡期间,heminodes 至少会失去一个末端分支,概率为 0.8。要删除的末端分支数量在 1 到 3 之间,概率相同。一个梅克尔细胞被删除的概率为 0.9。
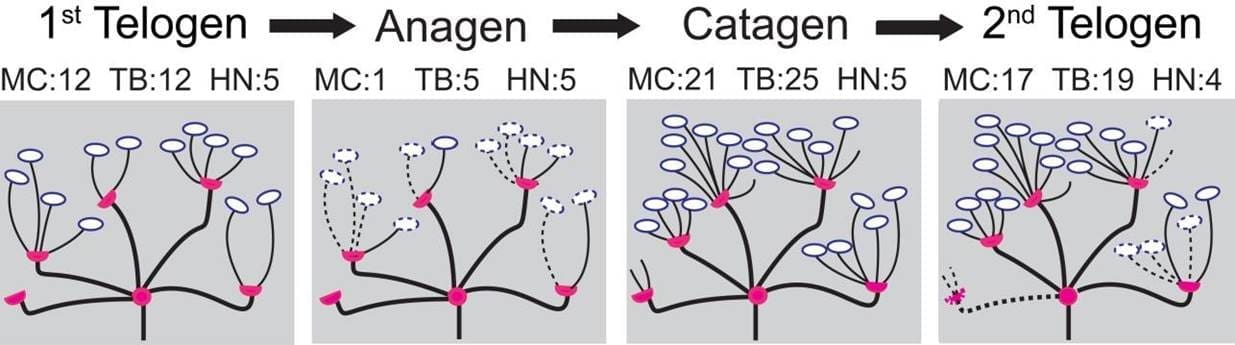
表 3 比较了从 20 个观察数据中得出的每个阶段梅克尔细胞和半结节的平均数量,以及从 20 次重复的 DES 模型中估算出的梅克尔细胞和半结节的平均数量。模型结果与实际数据相当相似,这表明基线模拟模型合理地代表了乔木系统(Banks,2000 年)。
表 2.模拟模型在毛发周期三个过渡阶段中每个阶段使用的参数值,其中 MC = 梅克尔细胞,TB = 末端分支,![]() 代表末端器官 x 在阶段s 中被移除/修剪的概率 x包括 MC、TB 和 HN。s 阶段包括 1T(第一次脱发)、A(生长期)、C(衰老期)和 2T(第二次脱发)。
代表末端器官 x 在阶段s 中被移除/修剪的概率 x包括 MC、TB 和 HN。s 阶段包括 1T(第一次脱发)、A(生长期)、C(衰老期)和 2T(第二次脱发)。
|
观测数据(平均值) |
模型输出(平均值 [95 % CI) |
|||
|
梅克尔细胞 |
半结节 |
梅克尔细胞 |
半结节 |
|
| 第一次脱发 |
13 |
5 |
13 [12.4, 17.3] |
5 [5.3, 5.5] |
| 衰老期 |
0 |
5 |
0 [0, 0.7] |
5 [5.2, 5.5] |
| 生长期 |
25 |
4 |
24 [24.1, 24.8] |
4 [3.9, 4.2] |
| 第二次脱发 |
15 |
3 |
14 [13.8, 14.2] |
3 [3.2, 3.9] |
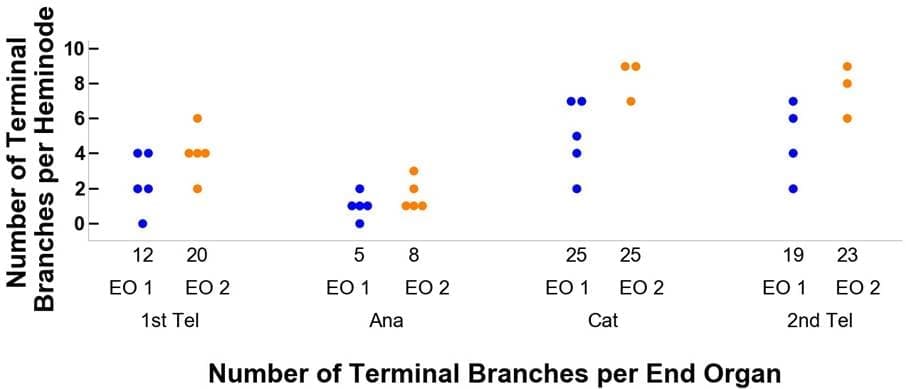
图 3 比较了观察数据和模拟数据的结果。仅依靠与毛发周期相关的线索,模型动力学产生的树干与实验观察结果非常一致。在每个毛发周期,每个末端器官的heminodes、末端分支和梅克尔细胞的分布以及变化模式与观察数据和模拟输出结果相似。通过模拟模型,我们还可以追踪毛发周期中每个末端器官的变化。例如,图 3 D、E 和 F 面板中的蓝色和橙色点分别代表两个不同的末梢器官。然而,由于观察数据缺乏可追溯性,我们没有在末端器官层面比较实验观察结果和模型输出结果之间的动态重塑行为。
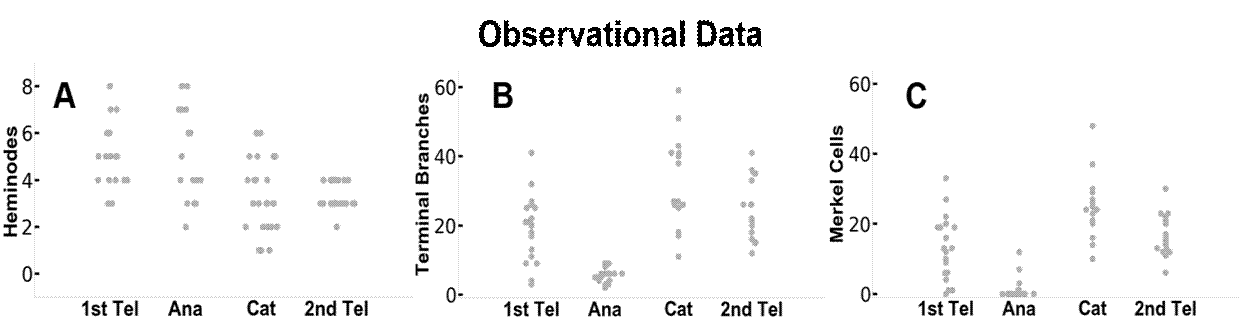
图 3.实际观察数据(A-C)与 DES 模型数据(D-F)之间的比较。 图中显示的是毛发周期四个阶段中每个阶段的神经节、末端神经分支和梅克尔细胞的相关数据。 在 D-F 面板中,分别用蓝色和橙色点来追踪毛发周期中两个不同阶段的末端器官。 这种样式也与图 4 中的数据相联系。
例如,在图 4A 中,末端器官 1(EO 1)开始时处于第一脱发期,有 12 个末端分枝和 5 个小节,末端分枝分布在小节上,每个小节有 4、4、2、2、0 个末端分枝。然后,当末梢器官 1 进入毛发周期的生长期阶段时,其末梢分枝数量减少到 5 个,这 5 个末梢分枝分布在 5 个小节上,对应关系为 2、1、1、1、0。 此外,请注意,在从生长期过渡到第二衰老期时,该末梢器官的小节数量从 5 个减少到 4 个。
为了了解哪些规则对末端器官重塑行为的影响更大,我们进行了一项敏感性分析。首先,研究了从第一衰老期到衰老期与末端分支和梅克尔细胞移除相关的规则。在过渡期,末端分支的删除取决于该末端分支是否有梅克尔细胞。对于未填充的末端分支,删除概率增加 0.1 会导致末端分支总数相差 20.5%,而填充末端分支的相同变化则导致相差 1%。此外,我们还评估了从白化期到第二衰老期修剪heminode的两条规则。分析结果表明,当无梅克尔细胞的细枝节的细枝节删除概率增加 0.05 时,细枝节的数量平均变化 6.7%。另一方面,对于有细胞的heminodes来说,同样的变化只会导致heminodes数量平均相差1.6%。从这些结果中,我们可以得出结论:末端器官成分的数量对人口较少的末端器官成分的删除政策更为敏感。
(A)
(B)
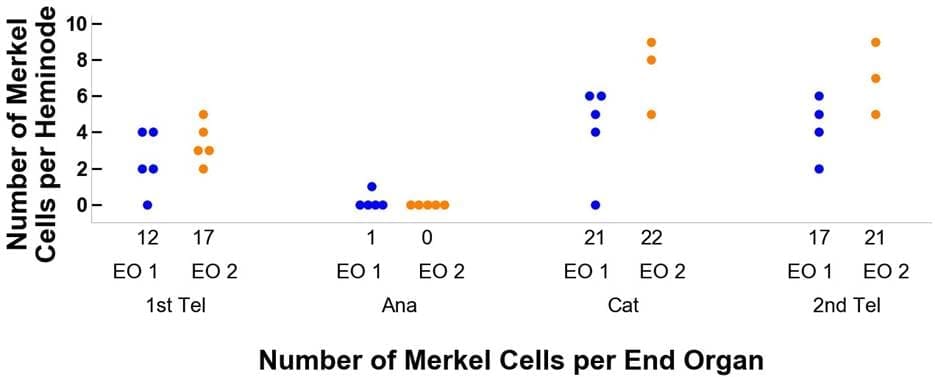
图 4.追踪两个末梢器官在毛发周期中从第一休止期、生长期、衰老期到第二休止期各阶段的流程。图 A 详细描述了末梢分支的变化,图 B 则与梅克尔细胞有关。 蓝色代表末梢器官 1(EO 1),橙色代表末梢器官 2(EO 2)。 例如,末梢器官 1 开始时有 12 个末梢分支和 12 个梅克尔细胞。然后进入毛发周期的生长期阶段,末端分枝的数量减少到 5 个,梅克尔细胞减少到 1 个,在 5 个细节上的分布分别为 2, 1, 1, 1, 0 和 1, 0, 0, 0。
讨论
如引言所述,本研究试图采用一种 DES 方法来确定毛发周期中心轴如何变化的原则,并评估梅克尔细胞和heminode动态的政策。我们选择了一种自下而上的方法,将末端器官、heminode和末端神经元分支作为不同的实体进行建模,从而完善了管理策略,并估算了在重塑过程中发挥作用的最佳参数。 该模型似乎很好地复制了观察到的数据,并在这一抽象水平上推测了一组可能驱动生物过程的概率值。模拟速率演化和合并规则的不同组合,使我们能够表现末端器官树干的动态,以便与自发毛发周期观察结果进行比较,并有助于填补观察研究中的局限性所留下的空白。例如,基于实体的自下而上的方法允许对毛发周期每个阶段的末端器官及其每个或每个组成部分进行详细的追踪,而这在观察研究中是受到限制的。此外,利用 DES 模型,我们还能确定毛发周期各阶段末端器官各组成部分的增减概率。 尽管如此,在建立 DES 模型时还是做了一些假设。首先,假设在后肢自发毛发周期数据集中观察到的平均值足够精确,可以制定合理的策略。其次,假定梅克尔细胞数量最少的heminodes最有可能被删除;另一方面,假定大梅克尔细胞簇中的梅克尔细胞更有可能被删除。最后,假设梅克尔细胞是随机添加到heminodes中的。
与在其他领域(如更广泛的医疗保健领域)的应用相比,DES 方法在生物研究问题上的应用还很有限(Hunt,2009 年)。本研究表明,DES 可作为生物研究中常用研究实践的一种有前途的附加方法。在转用小鼠模型之前测试潜在的生物规则,可以节省时间和金钱。使用 DES 模型进行各种假设情景测试和敏感性分析,还可以通过告知与突发动力学密切相关的关键因素,帮助设计额外的实验。 例如,该模型旨在提供参数实验功能。这种模型背后的动机是研究可能的混杂实验因素,如时间和末端器官条件,这些因素可能会影响末端器官的行为及其在毛发周期中的成分行为。具体来说,实验可以通过模拟这些不规则因素与基础模型概率规则的结合来确定所确定的概率规则的实际稳健性。
作者简历
HYOJUNG KANG是弗吉尼亚大学系统与信息工程系研究助理教授。她拥有宾夕法尼亚州立大学工业工程和运筹学博士学位。她的研究兴趣包括开发和应用混合仿真模型,为复杂系统(尤其是医疗保健系统)提供可实施的解决方案。她的电子邮件地址是 hkang@virginia.edu。
RACHEL L. ORLOWSKY 是弗吉尼亚大学系统与信息工程系和生物医学工程系的本科生。 她的兴趣是将计算建模应用于生物领域。她的电子邮件地址是 rlo4kc@virginia.edu。
GREGORY J. GERLING是弗吉尼亚大学系统与信息工程系和生物医学工程系的副教授。他在爱荷华大学获得博士学位,研究兴趣包括计算神经科学、触觉学、人为因素、人机交互。他的主要兴趣在于了解触觉系统的复杂性,这为我们的触觉感知和触觉设备的设计提供了依据。他的电子邮件地址是 gg7h@virginia.edu。